Jong-Bo Kim1,
Hyun Kang2 ![]()
For correspondence:- Hyun Kang Email: hkang@dankook.ac.kr Tel:+82415501452
Received: 6 November 2015 Accepted: 13 February 2016 Published: 31 March 2016
Citation: Kim J, Kang H. Ixeris dentata (Thunb) Nakai ethylacetate extract attenuates sterol regulatory element-binding proteins-1c via AMP-activated protein kinase activation. Trop J Pharm Res 2016; 15(3):453-459 doi: 10.4314/tjpr.v15i3.4
© 2016 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To investigate the molecular mechanisms underlying the role of Ixeris dentataa extract (IDE) in the prevention of high glucose-induced lipid accumulation in human HepG-2 hepatocytes.
Methods: IDE extract was prepared by maceration in ethyl acetate. Its fractionation was carried out by column chromatography. HepG-2 cells were pretreated with various concentration of IDE (0, 10, 20, 40 and 80 μg/mL) and then treated with serum-free medium with normal glucose (5 mM) for 1 h, followed by exposure to high glucose (30 mM D-glucose) for 24 h. Cell viability and cytotoxicity parameters were measured using lactate dehydrogenase (LDH) and MTT assay while triglyceride and total cholesterol levels were evaluated using respective enzymatic reagent kits. Protein ex
Results: IDE did not influence the cell viability (up to 200 μg/mL) and did not show any signs of cytotoxicity (up to 80 μg/mlL. IDE significantly attenuated lipid accumulation in human HepG2 hepatocytes when exposed to high glucose (30 mM D-glucose) in a dose-dependent manner (p < 0.05, 0.01 and 0.001 at 20, 40 and 80 μg/mL concentrations, respectively). Nile red staining showed that 10, 20, 40 and 80 μg/mL concentrations of IDE reduced lipid accumulation by 23.4, 34.8 (p < 0.05), 46.5 (p < 0.01) and 53.2 % (p < 0.001), respectively. The increased levels of triglycerides and total cholesterol were also attenuated by IDE (p < 0.001 at 80 μg/mL). Further, IDE attenuated the ex
Conclusion: The findings indicate that IDE exerts hypolipidemic effect by inhibiting lipid biosynthesis, mediated via AMPK signaling. This probably explains the extract’s beneficial effect in various inflammation disorders.
Introduction
Ixeris dentata (ID) is a well-known Korean traditional herb and has been commonly used in various diseases including respiratory and cardiovascular systems in Korea [1]. Since early times, both shoots and roots of ID have been used as plant-based foods such as kimchi and salad in Korea, and ID juice has been used to treat adult diseases [2]. More recently, secondary metabolites of ID have been known to possess anticancer, antistress, antiallergic, and antioxidant properties. Thus, ID is getting great interest as one of health functional medicinal plants [3]. However, the effect of an ethylacetate extract of ID and its molecular mechanism on lipid metabolism has not been elucidated.
Abnormal lipid and lipoprotein metabolism are known to result in dyslipidemia such as elevation of plasma cholesterol and/or TGs and decreased levels of high-density lipoprotein (HDL). Fatty liver disease is related to fat accumulation, high triacylglycerol, low high-density lipoprotein cholesterol (HDL-C), high low-density lipoprotein cholesterol (LDL-C), and insulin resistance [4].
A number of proteins associated with dyslipidemia have been reported to play important roles in lipogenesis, including fatty acid synthase (FAS), sterol regulatory element binding proteins (SREBPs) [2]. FAS are a multiple enzyme complex that catalyzes the synthesis of palmitate from acetyl-C glucose and malonyl-C glucose. Expression of FAS is regulated by multiple transcription factors. SREBPs, a transcription factor family, regulate the expression of enzymes for the synthesis of fatty acid, cholesterol, triacylglycerol, and phospholipids, suggesting that the protein family plays a central role in energy homeostasis via modulation of glycolysis, lipogenesis and adipogenesis.
Adenosine monophosphate-activated protein kinase (AMPK) is a serine threonine kinase comprising a heterotrimeric complex and regarded as an energy sensor in most tissues. AMPK regulates cellular lipid metabolism by increasing fatty acid oxidation and constitutive lipoprotein exocytosis [5]. In the liver, activated AMPK phosphorylates and inactivates the rate-limiting enzymes of lipogenesis such as ACC, suggesting that AMPK/ACC signaling plays a critical role in hepatic lipid homeostasis [6].
In previous study, ID contained approximately 4 mg/g of caffeic acid (3,4-dihydroxy cinamic acid) [2]. Approximately 20 types of sesquiterpene and other compounds have been isolated from ID. It has been reported that its main compounds are luteolin, luteolin 7-O-glucuronide, caffeic acid, chlorogenic acid, and guaiane sesquiterpene lactones [7,8]. Among these main compounds, inhibitory effects of luteolin, chlorogenic acid, and tectroside on allergic inflammation had already been reported. The major active components of ID are phenolic constituents [2,3].
This study aimed to examine whether ID extract (IDE) could suppress the hepatic lipid induced by high glucose and regulate lipid metabolism.
Methods
Plant material
The whole plant of I. dentata were collected on May 2014 at the herbarium were obtained from the local market, Seoul, South Korea. The fruits were authenticated by Professor Eun-Young Choi, a taxonomist, Konkuk University, Korea, and a voucher specimen (IDKU2014 was kept in the university herbarium for future reference. All reagents used in this study were of the highest commercial grade available.
Preparation of the ID extract
We employed a previous preparation method for this extract [9]. Briefly, 500 g of the fruit were ground in a mixer and defatted three times with three volumes of 80 % ethanol. The residue was extracted with absolute ethanol (EtOH) at 1:10 ratio (w/v) for 2 h on a heated mantle at 70 - 80 °C. The supernatant was filtered and concentrated in a rotatory evaporator at 50 °C. The ethanol extract obtained (180 g) was re-suspended in water:EtOH (9:1, v/v) and partitioned successively with n-hexane, ethyl acetate (EA) and n-butanol to obtain final yields of 19.4, 52 and 27.27 %, respectively. EA fraction of olive pulp fruit extract was dissolved in sterile distilled water and filtered on 0.22 μm filters before use.
Cell culture and treatment
Human HepG-2 hepatocytes obtained from the American Type Culture Collection (Rockville, MD) were cultured in Dulbecco’s modified Eagle’s medium (DMEM) containing normal glucose (5.5 mM d-glucose), supplemented with 10 % heat-inactivated foetal bovine serum (FBS; Invitrogen). Cells were cultured in a humidified 5% CO2 incubator at 37 °C in complete medium supplemented with 10 % FBS to 80 % confluence for use in the assays.
IDE was dissolved in dimethyl sulfoxide (DMSO) and added directly to serum-free culture medium. The final concentration of DMSO did not exceed 0.1 % (v/v) and did not affect cell viability nor produced cytotoxicity. A cell model of the high glucose induced accumulation of hepatic lipids was prepared by exposing HepG-2 cells to 30 mM of glucose for 24 h.
Measurement of cell viability and cytotoxicity
Cells were cultured at 37 oC in medium containing 10 % FBS at a density of 4 x 104 cells/500 µL in 48-well plates. After 24 h, the growth medium was replaced with serum-free medium and the cells were treated with different concentrations of IDE (10–100 µg/mL) or an equal volume of DMSO for 24 h at 37 oC. The culture medium was used in a lactate dehydrogenase (LDH) assay, with the absorbance at 490 nm measured using a microplate reader (Varioskan; Thermo Electron, Waltham, MA). Cytotoxicity was calculated based on the absorbance relative to that of the control. Cells were treated with MTT solution (final concentration, 0.5 mg/mL) for 1 h. The dark blue formazan crystals that formed in intact cells were solubilized with DMSO, and the absorbance at 570 nm was measured with a microplate reader. Cell viability was calculated based on the absorbance of the IDE-treated cells relative to that of control vehicle-treated cells.
Nile red stain
Human HepG2 hepatocytes were seeded in a 6-well plate (3 × 106 cells/well) and treated with glucose (30 mM) with indicated concentrations of IDE for 24 h. The cells were washed twice with PBS and fixed with 4 % formaldehyde in PBS for 1 h and then stained with 1 μg/mL Nile red for 30 min at room temperature. After staining, the distribution of lipid in cells was immediately analyzed by a FACScan flow cytometer (Becton Dickinson, Mountain View, CA, USA). Lipid-bound Nile red fluorescence was detected using inverted fluorescence microscopy.
Determination of triglyceride and total cholesterol levels
The intracellular triglyceride and total cholesterol contents were measured in cell lysates and expressed as micrograms of lipid per milligram of cellular protein. HEPG-2 cell were seeded in 100-mm culture dishes. Twenty-four hours after treatment, the cells were homogenized in 1 mL of PBS. The homogenates were extracted with 3 ml of chloroform and methanol (2:1, v/v). The mixture was vortexed vigorously, allowed to separate into two phases, and centrifuged (3000 rpm, 10 min, 4 oC). An aliquot of the organic phase was evaporated under nitrogen gas until dry. The resulting pellet was dissolved in PBS containing 1 % Triton X-100, and the protein concentration was measured. The intracellular triglyceride and total cholesterol contents were determined using an enzymatic reagent kit (Asan Pharm. Co., Seoul, Korea) at 550 and 500 nm, respectively, and normalized to the protein concentration.
Western blot analysis
Human HepG-2 were seeded in a 10 cm dish (1 × 106 cells/well) and treated with glucose (30 mM) at indicated concentrations of IDE for 24 h. The proteins of the cells were harvested in a cold radio immunoprecipitation assay (RIPA) buffer (1 % NP-40, 50 mM Tris-base, 0.1 % SDS, 0.5 % deoxycholic acid, 150 mM NaCl, pH 7.5). Equal amounts of protein samples were subjected to SDS–polyacrylamide gel electrophoresis and electro-transferred to nitrocellulose membranes (Millipore, Bedford, MA, USA). The membranes were blocked with 5 % nonfat milk powder with 0.1 % Tween-20 in TBS and then incubated with the first antibody at 4 °C overnight. Thereafter, membranes were washed three times with 0.1 % Tween-20 in PBS and incubated with the secondary antibody to anti-mouse horseradish peroxidase (GE Healthcare, Little Chalfont and Buckinghamshire, UK). Antibodies against AMPK and phospho-AMPK were purchased from Cell Signaling Technology (Beverly, MA, USA). FAS and SREBP-1c antibodies were obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Band detection was revealed by enhanced chemiluminescence using ECL Western blotting detection reagents and exposed ECL hyper film in FUJIFILM LAS-3000 (Tokyo, Japan).
Statistical analysis
Statistical analysis was performed using SAS program (SAS 9.2, SAS Institute Inc., USA). All measurements were carried out in quintuple Analysis of variance (ANOVA) was used to analyze the data followed by Duncan’s multiple range test and t-test. P < 0.05 was considered statically significant.
Results
Effect of IDE on HepG-2 cell viability
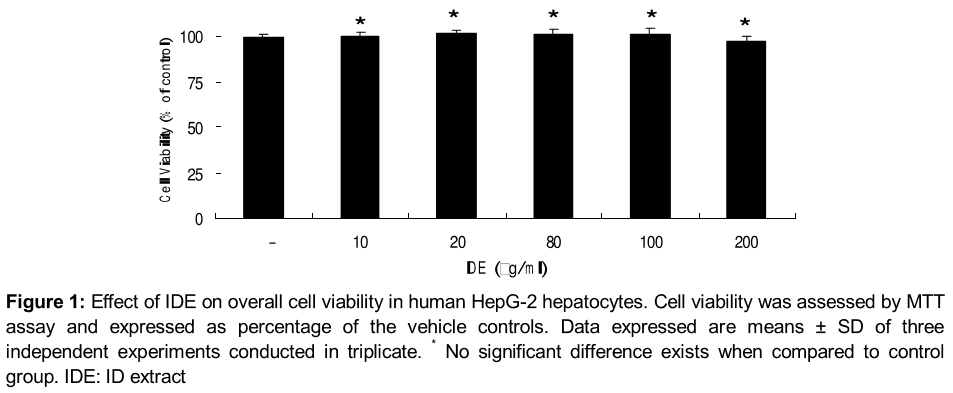
We first determined the concentration dependence of the cytotoxic effects of IDE by directly adding IDE to the culture medium at various concentrations (10 ~ 100 µg/ml) for 24 h, and the cell viability was measured using MTT assays ().
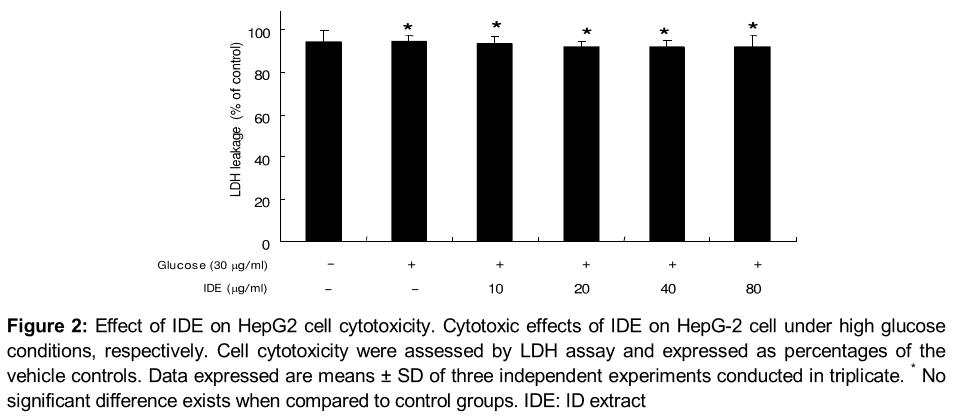
IDE also showed no signs of cytotoxicity at indicated concentrations (10 ~ 100 µg /ml) under normal glucose and high glucose conditions in HepG-2 cell (). Considering the results we used IDE in the range of 10 ~ 80 µg /ml, respectively for further experiments.
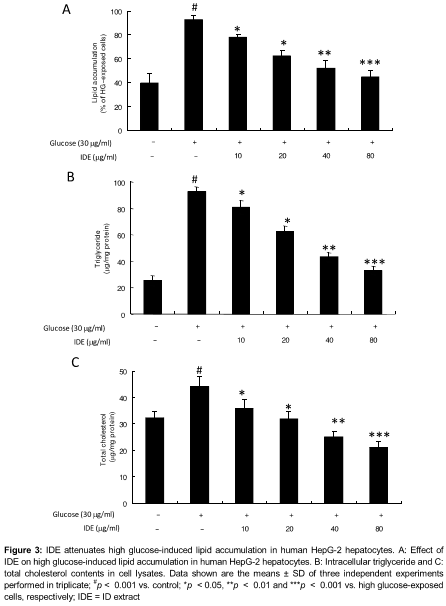
IDE attenuates lipid accumulation in human HepG-2 hepatocytes
To assess the inflammatory responses of IDE in vitro, HepG-2 cell were pretreated with various concentration of IDE (10, 20, 40 and 80 μg/mL) and treated with serum-free medium with normal glucose (5 mM) for 1 h, followed by exposure to high glucose (30 mM D-glucose) for 24 h. Nile red staining showed that 10, 20, 40, and 80 µg/mL concentrations of IDE reduced lipid accumulation by 23.4, 34.8, 46.5 and 53.2 %, respectively (A).
The effects of IDE on the triglyceride and total cholesterol levels showed that 20, 40, and 80 µg/mL of IDE reduced the triglyceride levels significantly, in the high glucose-treated cells by 38.9 %, 55.6 %, and 63.4 %, respectively (B). IDE at indicated concentrations also decreased the cholesterol content of HepG-2 cell incubated in high glucose for 24 h by 40.1, 52.1, and 61.2 %, at 20, 40, and 80 µg/mL concentrations, respectively (C). These results indicate that IDE can attenuate high glucose-mediated lipid accumulation in human HepG-2 hepatocytes.
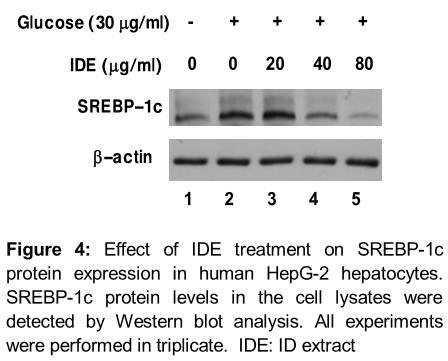
IDE inhibits lipid accumulation via SREBP-1c inactivation in human HepG-2 hepatocytes
We examined the status of SREBP-1c in high glucose-induced HepG-2 cell in the presence or absence of IDE at indicated concentrations. SREBP-1 protein levels were increased markedly in cells treated with high glucose (30 mM) for 12 h (, upper panel).
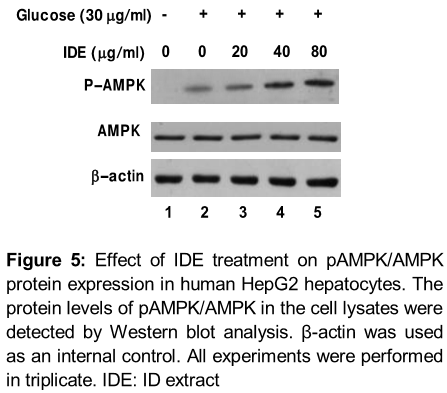
IDE promotes AMPK phosphorylation in human HepG-2 hepatocytes
The phosphorylation of AMPK was determined in HepG-2 cell under high glucose conditions by Western blotting. HepG-2 cell cultured in serum-free medium with normal glucose (5 mM) were pretreated with IDE (20, 40 and 80 µg/mL) for 1 h, followed by exposure to high glucose (30 mM) for an additional 2 h. Although the phosphorylation of AMPK was significantly suppressed in cells treated with high glucose (30 mM), IDE dose-dependently restored the phosphorylation of AMPK in IDE-pretreated cells ().
Discussion
The species, Ixeris dentata (ID) is a perennial herb of Asteraceae which was widely distributed and cultivated in Northeastern Asia [1]. The flowering time is from May to July and the flowers are ligulate and yellow [1]. This plant grows to height of about 25-50 cm [2]. The young sprouts of this species had been used as a famous bitter appetizing vegetable in Korea.
The plant has been used for the treatment of indigestion, pneumonia, hepatitis, contusion and tumor and various gastroenteric troubles in Korea [3]. According to recent research, ID has been proved to have a hypocholesterolaemic effect and an antioxidant activity, as well as ID is known to have aliphatics, triterpenoids and sesquiterpene glycosides in its composition [7,8]. Studies on the roots revealed the effects of an antioxidant, anti-mutagenicity and an anticancer. The methanol extract was most effective in the anticancer activity, inhibiting the growth of MG-63 cells by more than ninety percent [8]. Also, the effect of ID on the cardiovascular system in hyperlipidemic rats was examined [8] and also caused prevention of neurodegenerative diseases [9,10]. It has also been used for the treatment of allergic diseases as a folk therapy in Korea. The hepatic hypolipidemic mechanism of Ixeris dentata extract (IDE) showed high relevant expression of lipogenic enzyme, cholesterol biosynthesis and TG biosynthesis in HepG-2 cell. Taken together, these observations suggest that the neuro-inflammatory responses of IDE may require the AMPK signaling pathway. Activation of AMPK leads to regulation of a number of downstream targets involved in lipid metabolism [5]. The regulation of AMPK activation and the most effective constituent in IDE need to be elaborated.
Several natural products from plant origin have been suggested to be capable of preventing obesity via regulating hyperlipidemia and reducing adipose tissue mass, thus suppressing the occurrence of the metabolic, hepatic, and cardiovascular alterations associated with obesity [10,11]. Many studies have indicated that antioxidant phenolic compounds have been used in cellular and animal models for the treatment of obesity and neuro-inflammatory responses [13,14]. Polyphenols from blueberries and/or strawberries reduced the body weight and body fat of rats fed a high-fat diet [15]. Polyphenol rich antioxidants reduced hyperglycemia efficiently in type 2 diabetic mice via inducing AMPK expression in the liver and then resulted in significantly decreased liver and serum lipid contents [16].
The recent report showed that IDE possessed antioxidant properties [9, 10]. ID has been used for the treatment of mithridatism, calculous, indigestion, pneumonia, hepatitis, and tumors in Korea, China, and Japan [1]. It has been reported that ID has neuro-protective effects [10], anti-diabetic effects [11], protection effects against colitis and skin inflammation [2,3], and anti-allergic effects [10,11]. In this study, the effect of the IDE etylacetate extract on lipid metabolism via AMPK signaling was investigated in human HepG-2 hepatocytes. Earlier studies revealed that ID phenols possessed strong antioxidant compounds [18]. The ethyl acetate fraction present in ID might be responsible in exhibiting such potent antioxidant and anti-inflammatory effects.
Conclusion
The present study provides evidence that IDE plays a significant role in reducing HepG2 cellular lipid accumulation by mediating AMPK signaling, thereby inhibiting expression of SREBP-1c and further suppressing TG and cholesterol synthesis. The antioxidant potential of IDE might partly be responsible for this, suggesting an active role for IDE in preventing non-alcoholic fatty liver disease. Thus, the plant material has the potential to be developed to yield an anti-inflammatory agent.
Declarations
Acknowledgement
References
Archives


News Updates